Articol scris de Dina Mergeani
Pentru fiecare animal culoarea corpului (piele, par, pene) este un important mijloc de identificare, reprezentand si o strategie de supravietuire prin adaptarea la mediul inconjurator. Ceea ce ochiul percepe ca fiind culori ale porumbeilor sunt de fapt variate moduri de reflexie ale luminii determinate de aranjamentul pigmentilor (sau de absenta lor) de la nivelul penelor. Pigmentii sunt: derivati de caroten (cantaxantina – rosie, beta-caroten – galben) – la nivelul pielii picioarelor, derivati de melanina (eumelanina – neagra, feomelanima – rosie) – la nivelul penelor sau derivati de guanina (alb) si pteridina (galben) la nivelul ochilor.
Pigmentii melanici sunt produsi in anumite celule specializate numite melanocite. Acestea isi au originea in creasta neurala de unde migreaza sub forma de melanoblaste la nivelul ectodermului la inceputul vietii embrionare. Migratia, supravietuirea la locul de destinatie, proliferarea, transformarea im melanocite si patrunderea in ectoderm sunt procese controlate de doua perechi de gene care codifica doua perechi de proteine: c-kit protein si ligandul ei Steel (MCF), ednr-b si ligandul ei endotelina (edn-3). Modificarile acestor gene afecteaza procesele enumerate mai sus determinand aparitia la nivelul corpului a unor zone albe (fara melanocite) asociate in diferite modele de baltatura.
Odata ajunse la locul de destinatie, melanocitele produc pigmentii melanici la nivelul unor structuri numite melanozomi (eumelanozomi si feomelanozomi).
Feomelanozomii sunt mai mici si rotunzi, continand de trei ori mai putina tirozinaza decat eumelanozomii.
Eumelanozomii sunt mai mari, ovalari si contin si alte substante cum ar fi proteina p si enzima tyrp1.
Productia de pigmenti melanici incepe cu stimularea de catre alfa-MSH (hormon produs in hipofiza) a unui receptor de pe suprafata celulei numit MCR-1 care determina cresterea cantitatii de cAMP urmata de cresterea fosforilarii enzimei tirozinaza si a activitatii ei, consecutiv crescand productia de eumelanina. In ambele tipuri de melanozomi, sub influenta enzimei tirozinaza, aminoacidul tirozina este transformat in DOPA si apoi in DOPAquinone. In feomelanozomi, in prezenta aminoacidului cisteina DOPAquinone trece in cysteinDOPA care este transformata in feomelanina (de culoare rosie). In eumelanozomi DOPAquinone este transformata in DOPAchrome apoi in doi derivati (DHI, DHICA de culoare maron inchis) care, sub influenta enzimei tyrp-1, sunt convertiti in eumelanina (de culoare neagra). In anumite situatii aceste doua tipuri de pigment pot copolimeriza in variate proportii. Dupa ce pigmentii melanici sunt produsi in zona centrala a celulei ei sunt transferati keratinocitelor din jur si prin acestea penelor in crestere. Acest transfer se realizeaza cu ajutorul unei proteine transportoare (miozina 5) de-a lungul unor structuri pe baza de actina (microfilamente, microtubuli) din prelungirile melanocitelor. Acestea sunt asemanatoare dendritelor neuronale; melanocitele si neuronii avand o origine embriologica comuna. Cand ajung la nivelul membranei melanocitului, melanozomii elibereaza pigmentii melanici care vor fi incorporati in keratinocitele din jurul celulei si mai apoi in penele in crestere.
In concluzie, cei mai importanti factori ai melanogenezei sunt: tipul si activitatea tirozinazei, concentrantia de tirozina si concentratia de cisteina.
Genele mutante modifica procesul de melanogeneza prin producerea unor granule de pigment mai mici sau mai putine sau prin modificarea distributiei acestor granule: grupate sau distribuite uniform. Genele mutante recesive codifica proteine incapabile sa aiba o activitate normala, rezultatul actiunii lor fiind cel mai frecvent o modificare cantitativa. Genele dominante produc:
a. proteine normal functionale. Din acest punct de vedere genele Spread, Dirty, Sooty si cea care determina patternul solzat inchis la nivelul capacelor aripilor (T-pattern) pot fi considerate genele care produc enzimele cu o activitate normala.
b. blocanti ai unor sinteze proteice prin legarea competitiva de locusul de actiune al proteinelor normale. In al doilea caz aceste gene produc uzual modificari calitative. Astfel gena corespondenta wild-type situata pe locusul Smoky impiedica dispersia granulelor de pigment la nivelul zonelor aproape albe ale unui porumbel Wild-type: dungile din partea laterala a penelor laterale ale cozii, partea dinspre interior (torace) a penelor de la nivelul capacelor aripilor si spatele inferior. Ea reprezinta o modificare adaptativa, porumbeii cu spatele inferior alb fiind atacati mai rar de pradatori.
In acest fel putem spune ca, din punct de vedere enzimatic, un porumbel S//S, C*T//C*T, V/V, So//So, sy//sy este tipul normal de porumbel.
Mutatiile cele mai cunoscute sunt:
1. Mutatii la nivelul genei care codifica MCR-1 (extension locus). Mutatiile produc variante ale acestui receptor de care alfa-MSH nu se poate lega, rezultand o enzima tirozinaza cu o activitate scazuta si preponderenta productiei de feomelanina.
La porumbei avem trei alele ale acestui locus:
– gena normala (wild-type) ce duce la productia normala de eumelanina;
– gena ember (e*Em) care determina o scadere a productiei de eumelanina;
– gena recessive red (e) care produce o scadere marcata (pana aproape de zero) a productiei de eumelanina.
2. Mutatii la nivelul genei care codifica enzima tirozinaza (albino locus). Aceste mutatii produc enzime total sau aproape total nefunctionale determinand melanocite aparent normale morfologic dar care nu sunt capabile sa produca pigmenti melanici.
La porumbei avem trei alele ale acestui locus:
– gena normala cu o enzima functionala (wild-type);
– gena albino-homer (al*H) care produce o enzima aproape total inactiva, porumbeii prezentand urme de pigment la nivelul ochilor;
– gena albino (al) care produce o enzima total inactiva, porumbeii avand ochii complet rosii.
3. Mutatii la nivelul genei care codifica proteina p (pink eye dilute locus). Aceasta proteina, care functioneaza ca un regulator al pH-ului din eumelanozomi, poate determina variatii ale activitatii tirozinazei.
Prezinta doua alele:
– gena normala, care determina un pH acid cu o enzima tirozinaza activa si cu o cantitate normala de eumelanina;
– gena pink eye dilute (pd) care produce o proteina p modificata ce determina un pH neutru, consecutiv scazand si activitatea tirozinei si cantitatea de eumelanina. Dat fiind ca proteina p nu este prezenta in feomelanozomi, modificarea ei nu afecteaza productia de feomelanina.
4. Mutatii care codifica enzima tyrp-1 (brown locus).Alela normala produce o enzima activa care duce la o productie normala de eumelanina (de culoare neagra), gena tyrp-1(b) producand o enzima cu o activitate mai redusa care nu poate transforma precursorii DHI, DHICA (de culoare maroniu inchis) in eumelanina. Desi intre culoarea maroniu-ciocolatiu si cea albastru-negru sunt diferente de oxidare a pigmentilor, la prima culoare eumelanozomii fiind mai mici si mai rotunzi, s-a emis si ipoteza ca aceasta gena ar putea sta la originea culorii maroniu-ciocolatiu la porumbei.
5. Mutatii care codifica proteina miozina 5 (myo-5) situate pe locusul Dilute. Aceste mutatii produc proteine cu activitate scazuta care transporta doar o parte din melanozomi prin dendrite spre membrana celulara avand ca rezulta o scadere a cantitatii de pigment incorporate de keratinocitele din jur. Aceaste modificari afecteaza ambele tipuri de pigment scazand intensitatea culorii. La porumbei genele acestui locus sunt: wild-type (intense), pale (d*p, cu o scadere de ~25% a cantitatii de pigment), dilute (d, cu o scadere de ~50% a cantitatii de pigment), ecru (d*ec, contitatea de pigment fiind foarte putina) si, probabil, blind dilute.
6. Despre celelalte mutatii se presupune ca:
– gena Spread determina cresterea cantitatii de tirozina si a activitatii tirozinazei. De fapt Spread creste concentratia de eumelanina de aproximativ 6 ori si pe fond albastru-negru si pe fond rosu-cenusiu fara sa modifice concentratia de feomelanina;
– genele Dirty, Sooty, Smoky si pentru variantele de solzati (Checkers) determina cresterea cantitatii de tirozina, Dirty actionand ca o gena Spread mai putin eficienta, Sooty si genele Patternului cresc cantitatea totala de pigment (indiferent daca acesta este negru sau rosu) iar Smoky creste in mai mare masura cantitatea de pigment rosu;
– genele din grupul celor care produc variate tipuri de bronz, Indigo, Recessive opal, Ash-red (si altele) produc o scadere a tirozinei dar mai ales a tirozinazei, ducand la aparitia unor zone de bronz in locul zonelor negre, o arie cu o nuanta de bronz fiind o arie in care exista o mixtura de eumelanina si feomelanina. De fapt gena Ash-red scade concentratia de eumelanina de aproape 12 ori fara sa modifice concentratia de feomelanina, actionand ca un blocant al productiei de eumelanina iar gena Indigo are un efect similar dar mai slab;
– genele din grupul Grizzle si din grupul Almond produc o scadere mai ales a tirozinei dar si a tirozinazei, ducand la prezenta unei zone de bronz intre aria colorata si cea alba a unei pene; – gena Penciled scade cantitatea de tirozina – daca nu este asociata una din genele pentru bronz nu exista o zona cu bronz intre zona alba si cea neagra a penei. Orice referire la acest material se va face mentionand autorul, numele articolului si adresa site-ului unde este prezentat.

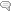
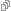

 Publicat de Andrei Bondar
Publicat de Andrei Bondar 
 Gusat slovac (Expo Timisoara 2006 // D. Mergeani)
Gusat slovac (Expo Timisoara 2006 // D. Mergeani) Porumbel Modena (Expo Timisoara 2006 // D. Mergeani) (Ts1//?, V//?)
Porumbel Modena (Expo Timisoara 2006 // D. Mergeani) (Ts1//?, V//?) b. Porumbeii cu penajul matasos au un zbor greoi deoarece lama penei este destramata. Acest caracter fenotipic poate fi urmarea actiunii independente a mai multor gene:
b. Porumbeii cu penajul matasos au un zbor greoi deoarece lama penei este destramata. Acest caracter fenotipic poate fi urmarea actiunii independente a mai multor gene:




















































